
A half-century ago, the great pioneers of ethology (e.g. Tinbergen 1951) and neuroscience (e.g. Hebb 1949) recognized that the need to bridge the gap between the ‘ethologist’ and the ‘physiologist’ was integral toward achieving a clear understanding of the behavior of the ‘intact animal’. Today, as integrative approaches in biology steadily gain increasing popularity and as vast technological advances have been made, perhaps ironically, the distinctions in the study of evolution Tinbergen envisioned (including describing the course of evolution, the genetics proper of behavior, and the directional changes under the influence of natural selection) have begun to coalesce. A multi-level approach toward accounting for genetic, brain, and behavioral factors offers the opportunity to gain a better understanding of mechanisms of behavior, and offer insight into the causes of both the evolution and behavior of Tinbergen’s ‘intact animal’.
Our lab is unified in our quest to better understand social behavior. Sociality is one of the most intriguing topics in the study of animal behavior because it is so pervasive. Work in our lab can be generally categorized under the following questions.
1) What is the “Social Brain”?
Social behavior is found in nearly every known animal, although the degree of sociality can vary profoundly in terms of form and frequency. If we are to ever gain an accurate understanding of social behavior, we must first 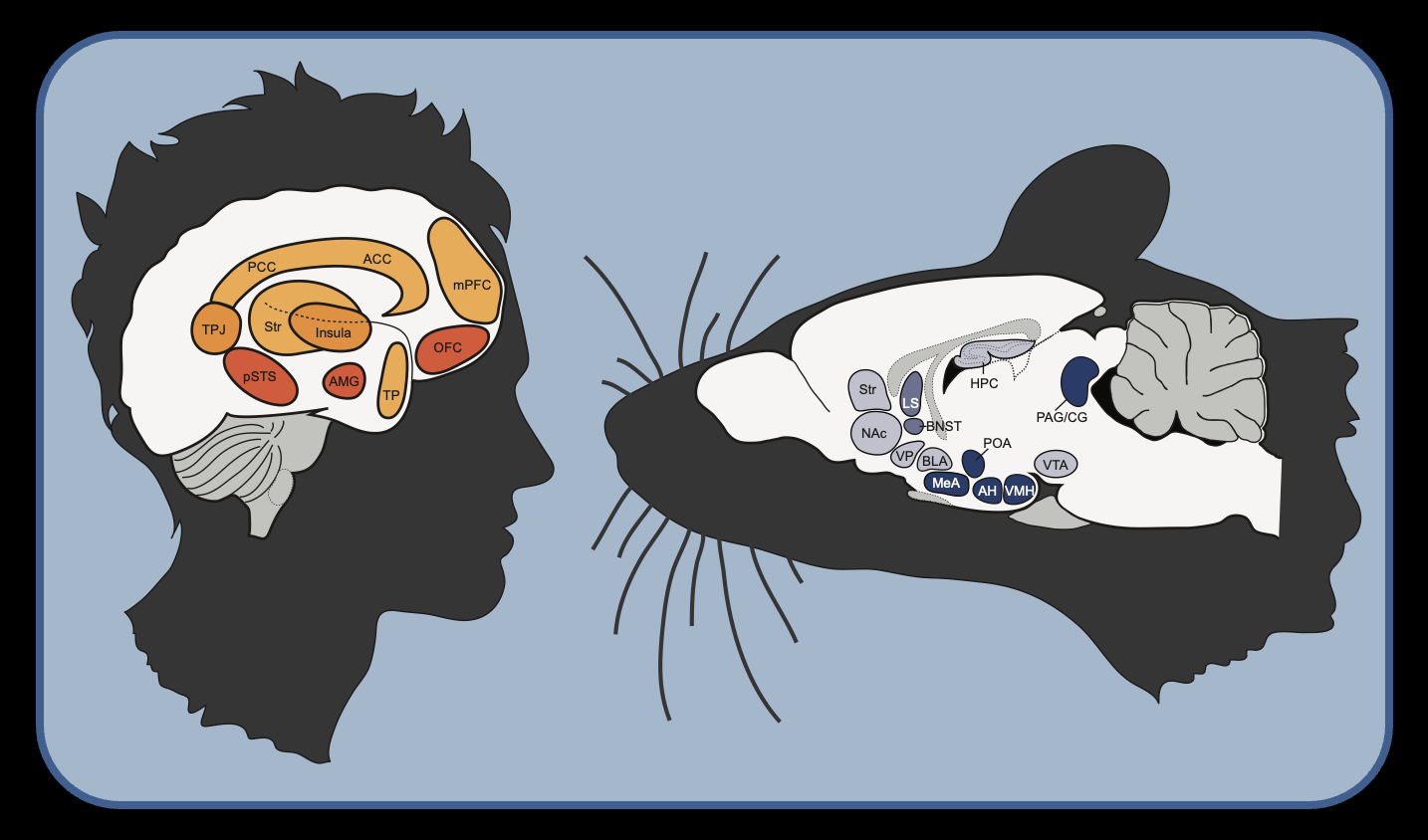 recognize the many different forms that social behavior takes. Indeed, social behavior can be subdivided into relatively distinct but interrelated categories of behaviors (like affiliation, aggression, approach, social grouping, consolation, mating, nurturing, play, etc.). Each of these complex behaviors involves many different behavioral elements and interactions. The neural control of social behavior is inherently even more complex than the behaviors themselves.
recognize the many different forms that social behavior takes. Indeed, social behavior can be subdivided into relatively distinct but interrelated categories of behaviors (like affiliation, aggression, approach, social grouping, consolation, mating, nurturing, play, etc.). Each of these complex behaviors involves many different behavioral elements and interactions. The neural control of social behavior is inherently even more complex than the behaviors themselves.
For as long as people have studied social behavior, an underlying goal has been to understand the mechanisms that govern them. Neuroscientists with different backgrounds and expertise have attempted to define the core neural mechanisms that underlie social behavior, but defining a ‘social brain’ is as complicated as the behaviors that are presumably under its control. Yet, our field often suffers from taking an overly simplistic view of brain-behavior relationships, and commonly promotes the idea that brain-behavior relationships can be satisfactorily explained by a one-to-one mapping of neural circuits to specific behaviors. Despite the tremendous importance of understanding the ‘social brain’, it is naïve to assume that all aspects of social behavior are under the control of the same neural subcircuits. Notably, some elements of the social brain are shared across different social behaviors, and presumably represent points of communication that offer powerful opportunities to understand the depth of the functional social brain. Several attempts have been made to characterize the social brain, and the most dominant views are responsible for tremendous progress toward understanding social behavior. Nevertheless, determining how and where the brain processes and shapes behavior in response to social factors is of great importance if we are to ever truly understand the nature and universality of social behaviors.
2) How do early life social experiences (both positive and negative) – and paternal care in particular – impact offspring development and adult brain-behavior phenotypes.
Interaction between parents and offspring is the cornerstone of emotional development. Early social experience impacts behavior from the cradle to the grave. Our work investigates the po tential for social experiences to enhance or impede development of the brain, social behavior, and cognition. We are exploiting the rich potential for prairie voles to serve as the gold-standard model for paternal (and maternal) care. We have found that manipulating the life history of offspring in the postnatal environment (e.g., rearing in the presence or absence of fathers and/or same-aged peers), alters socio-spatial cognition, affiliation, social approach, and impacts OTR and V1aR density in key regions of the ‘social brain’. The nonapeptide systems are highly dynamic in the developing postnatal brain and influenced by social and environmental complexity, but largely stable in adults – particularly in pregnant females. We have identified a putative pathway whereby variation in paternal care leads to alterations in DNA methylation within the vasopressin receptor 1a (V1aR) gene and V1aR gene expression in the lateral septum. These differences are concomitant with changes in social approach, a behavior closely associated with septal V1aR function. We have also found that the presence or absence of the father impacts the parental care of mothers. All of this work points to the fact that fathers play a crucial role in the social and neural stability of offspring.
tential for social experiences to enhance or impede development of the brain, social behavior, and cognition. We are exploiting the rich potential for prairie voles to serve as the gold-standard model for paternal (and maternal) care. We have found that manipulating the life history of offspring in the postnatal environment (e.g., rearing in the presence or absence of fathers and/or same-aged peers), alters socio-spatial cognition, affiliation, social approach, and impacts OTR and V1aR density in key regions of the ‘social brain’. The nonapeptide systems are highly dynamic in the developing postnatal brain and influenced by social and environmental complexity, but largely stable in adults – particularly in pregnant females. We have identified a putative pathway whereby variation in paternal care leads to alterations in DNA methylation within the vasopressin receptor 1a (V1aR) gene and V1aR gene expression in the lateral septum. These differences are concomitant with changes in social approach, a behavior closely associated with septal V1aR function. We have also found that the presence or absence of the father impacts the parental care of mothers. All of this work points to the fact that fathers play a crucial role in the social and neural stability of offspring.
3) What are the neural mechanisms subserving mating tactics, social monogamy, sexual fidelity, and reproductive decision-making?
The maintenance of healthy committed relationships has significant impacts on the physical and mental health of humans. Our knowledge about the neurobiology of pair bonding has been advanced through the development of the prairie vole model for attachment and love. However most of this work has centered on laboratory work that ignores the individual variation in sexual fidelity. Our research program has been unique in its aim to account for  individual variation in neural mechanisms governing sexual fidelity in real-world contexts. Several of our papers have detailed the distinctions in behavior and the brain of males and females that engage in monogamous and non-monogamous mating tactics in the field. This work has demonstrated that variation in oxytocin and vasopressin receptors (OTR and V1aR) in areas necessary and sufficient for establishing pairbonds usually do not vary across individuals, but that structures associated with social and spatial memory predict reproductive success in paired and single animals. We have also demonstrated that DREADD activation of the lateral septum facilitates prosociality but does not affect selective affiliation with a partner.
individual variation in neural mechanisms governing sexual fidelity in real-world contexts. Several of our papers have detailed the distinctions in behavior and the brain of males and females that engage in monogamous and non-monogamous mating tactics in the field. This work has demonstrated that variation in oxytocin and vasopressin receptors (OTR and V1aR) in areas necessary and sufficient for establishing pairbonds usually do not vary across individuals, but that structures associated with social and spatial memory predict reproductive success in paired and single animals. We have also demonstrated that DREADD activation of the lateral septum facilitates prosociality but does not affect selective affiliation with a partner.
This work has led to our hypotheses that 1) monogamy is inherently a socio-spatial task, and 2) that individual variation in sociability contributes to the variation observed in sexual fidelity and promiscuity. The former hypothesis is based on the idea that mating tactics require an accounting of the spatial distribution of conspecifics in the social landscape (i.e., knowing who and where potential mating partners or competitors can be found). The latter hypothesis is based on the idea that sociable individuals are more likely to engage in indiscriminate prosocial behaviors, which creates opportunities to engage in extra-pair copulations. This work has broadened our current understanding of the neurobiology of social behavior and social attachment, with specific emphasis on monogamy-related behaviors (e.g., selective affiliation, territorial aggression, parental care). We also challenge existing ideas of what contributes to ‘monogamy’, and argue against biases from Western cultures that conflate sexual fidelity and social bonds. Indeed, a rigorous understanding of social behavior requires a disentangling of falsely intertwined concepts.
4) Understanding the interface between reproductive physiology and social communication at the genomic, social, and reproductive substrates of odor discrimination*.
The ability to locate, identify, and select mates most suited to reproduction is a critical component of individual fitness and is fundamental to all forms of life. In many vertebrate taxa, mate discrimination for the purposes of reproduction is mediated by chemical communication, placing the chemical senses in a position of significant biological importance. Despite this, we have a relatively poor and limited understanding of what the genetic and biological substrates that facilitate detection, identification, and evaluation of potential mates are. Rodents have provided much of the foundational work in the area of chemosensory mechanisms. However, laboratory populations of mice and rats, the preferred mammalian models for such work, are usually genetically identical and have not been exposed to a natural environment for many generations, limiting their usefulness for research on interactions within natural environments or for understanding selective pressures driving the evolution of chemosensory systems. We areexamining the genomic and physiological factors contributing to chemosensory systems in an exciting and emerging model species of rodent that allows a rare opportunity to study mechanisms of chemical communication and mate choice in a unique reproductive system.
The African giant pouched rat, Cricetomys ansorgei, provides an exquisite opportunity to study the dynamics of chemical communication. Natural populations of pouched rats are genetically diverse and have been exposed to unique social and environmental selective pressures unlike traditional laboratory rodents. Notably, this species possesses extremely powerful chemosensory abilities, even by comparison to other rodents. The acute chemical senses of the pouched rat are powerful enough to detect explosives, and as a result, pouched rats are frequently touted for their significance as a ‘biodetector’. The natural and superb ability of this species to detect explosives might seem peculiar, but it reflects the deep evolutionary roots aimed at mate attraction and discrimination. This is because TNT is closely associated with a class of chemicals known as nitromusks, frequently used in the perfume industry and naturally used to advertise to and attract animals of the opposite sex. Therefore, the advanced and highly sensitive biodetection abilities of pouched rats to detect chemicals such as TNT very likely evolved from the ability to detect social signals related to mate attraction (i.e., musk and other pheromones). By developing a clearer understanding of the mechanisms that govern olfactory mate discrimination, we can also gain a better understanding of the capacity and limits of rodent biodetection, while elucidating one of the most fundamental forms of social communication.
*[This project is no longer active, but you can visit our animals and their offspring at several zoos across the country including: Bronx Zoo (NY), Chehaw Zoo (GA), Cincinnati Zoo (OH), Dallas Zoo (TX), Hogle Zoo (UT), Little Rock Zoo (AR), Memphis Zoo (TN), Point Defiance Zoo (WA), Roger Williams Zoo (RI), Rosemond Gifford Zoo (NY), Staten Island Zoo (NY), and The Wild Animal Park (NY)]
Our work with prairie voles has been supported by generous funding from NIH (NICHD) and NSF (IOS, Neural Systems).
Our work with African giant pouched rats was supported by generous funding from the Department of Defense.